RANKL/RANK signaling (WP2018)
Homo sapiens

{kind=link}
{kind=link}
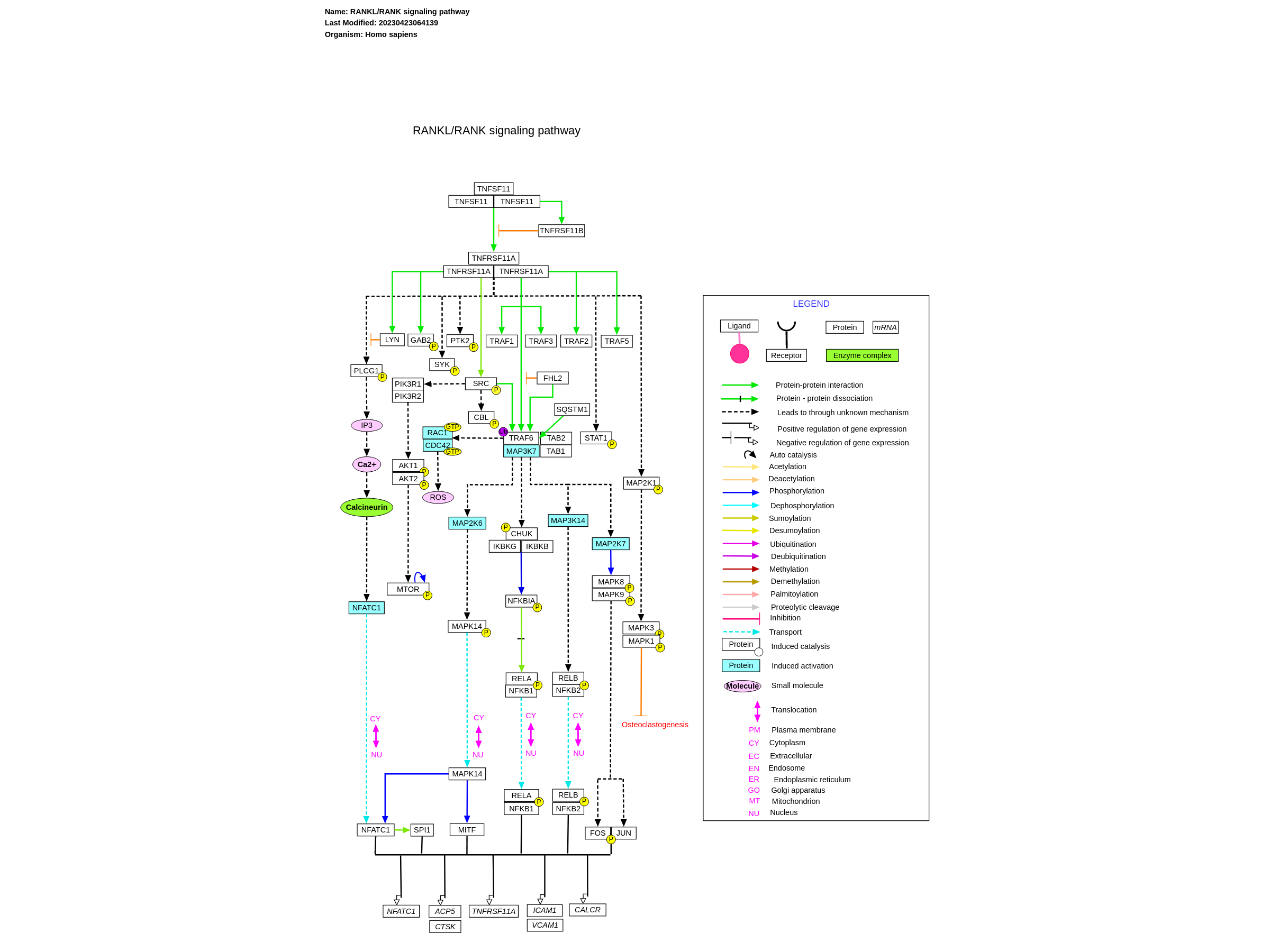
RANKL (Receptor activator of nuclear factor-kappa B ligand), RANK (Receptor activator of nuclear factor-kappa B) and the natural decoy receptor of RANKL, OPG (Osteoprotegerin) are three important molecules identified to play a major role in osteoclastogenesis and bone remodelling. They are members of the tumor necrosis factor (TNF) superfamily. OPG was the first molecule to be discovered and proved to inhibit osteoclastogenesis both in vivo and in vitro. Unlike other members of TNF family, OPG lack a transmembrane domain and is secreted as a soluble protein by the cell. RANKL is the only known physiological agonist for its receptor, RANK. Genetic experiments have shown that mice lacking either rankl or rank suffer from severe osteoporosis and defective tooth eruption due to complete lack of osteoclasts. On the contrary, mice deficient of OPG shows osteoporosis due to increased number of osteoclasts. Binding of RANKL to RANK triggers downstream signaling events that leads to the activation of osteoclasts and controlling of lineage commitment. RANKL/RANK signaling is essential for skeletal homoeostasis and its interference leads to inhibition of bone resorption resulting in bone diseases including osteoporosis osteopetrosis and rheumatoid arthritis. RANK being a member of TNF family does not possess any kinase activity. It recruits adaptor molecules to transduce the signal after ligand binding. These adaptor molecules are called TNFR-associated factors or TRAF’s that binds to different regions in the cytoplasmic tail of the TNF family receptors and transduces the signal downstream. TRAF6 is the main adaptor molecule which activates NF-κB pathway downstream of RANKL signaling which is required for osteoclastogenesis and osteoclast activation. TRAF6 mutant mice have shown a partial block in osteoclastogenesis and defective activation of mature osteoclasts. Mice lacking NF-κB p50 and p52 proteins have been shown to be osteopetrotic. Catalytic subunits, IκB kinase α and IκB kinase β and the non-catalytic subunit IKKγ (also called NEMO) are also essential for RANKL-RANK signaling and osteoclastogenesis. IKKγ is required for osteoclastogenesis induced by RANKL in mice both in vivo and in vitro whereas IKKα was shown to be required in mice only in in vitro. Several mitogen activated protein kinases (MAPK’s) have been shown to be activated downstream of RANK. Studies have shown that pharmacological inhibition of p38 MAPK’s blocked RANKL induced osteoclast differentiation. JNK1/2, its upstream kinase MKK7 and c-Jun have also been shown by genetic experiments to be essential for RANKL induced osteoclastogenesis. MAPK1 and MAPK3 phosphorylation was also shown to be dispensable for RANKL mediated osteoclast differentiation in vitro, but another report also show that specific inhibitors to MEK increased RANKL induced osteoclastogenesis suggesting a cross talk between p38 and ERK signaling pathways. NFATc1 is an essential downstream target of RANK. Ca2+ oscillations induced by RANKL activated NFATc1 resulting in terminal differentiation of osteoclasts through the Ca2+- dependent calcineurin pathway. NFATc1 translocates to the nucleus where it interacts with other transcription factors leading to the activation of transcription of genes including ACP5, CTSK, TNFRSF11A and NFATc1 under RANKL stimulation. TRAF6 and c-Src interacts with each other and with RANK upon stimulation with RANKL. This interaction increases the kinase activity of c-Src leading to the tyrosine phosphorylation of downstream molecules such as c-Cbl and activation of Akt/PKB which in turn requires the PI3-Kinase activity. Genetic experiments have shown that c-Src is very important in osteoclastogenesis. In addition to these pathways, aPKC/p62 signaling is also reported to be essential for osteoclastogenesis. Apart from their role in osteoclast differentiation and function, RANKL-RANK signaling is also required for development of lymph node and lactating mammary glands in mice and in the establishment of thymic microenvironment. Please access this pathway at NetSlim database. If you use this pathway, please cite following paper: Raju, R., Balakrishnan, L., Nanjappa, V., Bhattacharjee, M., Getnet, D., Muthusamy, B., Thomas, J. K., Sharma, J., Rahiman, B. A., Harsha, H. C., Shankar, S., Prasad, T. S. K., Mohan, S. S., Bader, G. D., Wani, M. R. and Pandey, A. (2011). A comprehensive manually curated reaction map of RANKL/RANK signaling pathway. Database (Oxford). 2011, bar021.
For a description of pathway objects, see the WikiPathways Legend.
Authors
Jyoti Sharma , Kristina Hanspers , NetPath , Daniela Digles , Egon Willighagen , Zahra Roudbari , Chris Evelo , Martina Summer-Kutmon , Alex Pico , Denise Slenter , and Eric WeitzActivity

Discuss this pathway
Check for ongoing discussions or start your own.
Cited In
- Hematopoietic stem-cell senescence and myocardial repair - Coronary artery disease genotype/phenotype analysis of post-MI myocardial regeneration response induced by CABG/CD133+ bone marrow hematopoietic stem cell treatment in RCT PERFECT Phase 3 (2020).
- MicroRNAs as potential biomarkers for doxorubicin-induced cardiotoxicity.
- A Practical Strategy for Exploring the Pharmacological Mechanism of Luteolin Against COVID-19/Asthma Comorbidity: Findings of System Pharmacology and Bioinformatics Analysis (2024).
Are you planning to include this pathway in your next publication? See How to Cite and add a link here to your paper once it's online.
Organisms
Homo sapiensCommunities
Annotations
Pathway Ontology
signaling pathway pertinent to development| Label | Type | Compact URI | Comment |
|---|---|---|---|
| IP3 | Metabolite | hmdb:HMDB0001498 | |
| ROS | Metabolite | chebi:26523 | |
| NFKB1 | Protein | ncbigene:4790 | |
| FOS | Protein | ncbigene:2353 | |
| RELB | Protein | ncbigene:5971 | |
| RAC1 | Protein | ncbigene:5879 | |
| MAP2K1 | Protein | ncbigene:5604 | RANKL stimulation induces MEK1 phosphorylation in RAW264.7 cells |
| FHL2 | Protein | ncbigene:2274 | |
| MITF | Protein | ncbigene:4286 | |
| LYN | Protein | ncbigene:4067 | |
| MAP2K6 | Protein | ncbigene:5608 | |
| SYK | Protein | ncbigene:6850 | RANKL stimulation induces SYK phosphorylation at Tyr-323 and Tyr-352 in mouse derived bone marrow cells and RAW264.7 cells |
| TNFRSF11B | Protein | ncbigene:4982 | |
| MAP2K7 | Protein | ncbigene:5609 | |
| TRAF6 | Protein | ncbigene:7189 | TRAF6 undergoes polyubiquitination upon stimulation with RANKL which is further enhanced by IFN-gamma in mouse macrophage cell line RAW 264.7 and in mouse bone marrow derived macrophages. |
| TNFSF11 | Protein | ncbigene:8600 | |
| GAB2 | Protein | ncbigene:9846 | |
| TNFRSF11A | Protein | ncbigene:8792 | |
| TRAF2 | Protein | ncbigene:7186 | |
| TRAF3 | Protein | ncbigene:7187 | |
| TRAF5 | Protein | ncbigene:7188 | |
| TRAF1 | Protein | ncbigene:7185 | |
| PIK3R1 | Protein | ncbigene:5295 | |
| NFATC1 | Protein | ncbigene:4772 | |
| PIK3R2 | Protein | ncbigene:5296 | |
| STAT1 | Protein | ncbigene:6772 | RANKL stimulation induces STAT1 phosphorylation at Ser-727 in mouse pre-osteoclasts and bone marrow macrophages |
| IKBKG | Protein | ncbigene:8517 | |
| IKBKB | Protein | ncbigene:3551 | |
| AKT1 | Protein | ncbigene:207 | AKT1 undergoes induced phosphorylation upon stimulation with RANKL at Thr-308 and Ser-473 in mouse bone marrow derived macrophages and in maouse macrophage cell line, RAW264.7. |
| NFKBIA | Protein | ncbigene:4792 | NFKBIA undergoes induced phosphorylation at Ser-32 in mouse bone marrow macrophages and in RAW264.7 cells. |
| MAPK9 | Protein | ncbigene:5601 | RANKL induces MAPK9 phosphorylation at Thr-183 and Tyr-185 in mouse bone marrow derived macrophages. |
| MAP3K7 | Protein | ncbigene:6885 | |
| PLCG1 | Protein | ncbigene:5335 | RANKL stimulation induces PLCG1 phosphorylation in bone marrow derived macrophages and RAW264.7 cells. |
| JUN | Protein | ncbigene:3725 | JUN undergoes induced phosphorylatin at Ser-63 and Ser-73 in mouse bone marrow derived macrophages, preosteoclasts and 293 cells. Amino acid residues 340-421 of TNFRSF11A are required for its activation |
| PTK2 | Protein | ncbigene:5747 | RANKL stimulation induces PTK2 phosphorylation in human umbilical vein endothelial (HUVEC) cells and RAW264.7 cells |
| RELA | Protein | ncbigene:5970 | RELA undergoes induced phosphorylation at Ser-276 and Ser-536 upon stimulation with RANKL in mouse bone marrow macrophages. |
| AKT2 | Protein | ncbigene:208 | |
| MAPK8 | Protein | ncbigene:5599 | RANKL induces MAPK8 phosphorylation at Ser 183 and Tyr-185 in mouse bone marrow macrophages and preosteoclasts |
| MAPK3 | Protein | ncbigene:5595 | MAPK3 undergoes induced phosphorylation upon stimulation with RANKL at Thr-202 and Tyr-204 in mouse macrophage cell line, RAW264.7, mouse bone marrow derived macrophages, osteoclasts and dendritic cells. |
| MAPK1 | Protein | ncbigene:5594 | MAPK1 undergoes induced phosphorylation upon RANKL stimulation in mouse macrophage cell line, RAW264.7, mouse bone marrow derived macrophages, osteoclasts and in dendritic cells. |
| MAPK14 | Protein | ncbigene:1432 | MAPK14 undergoes phosphorylation at Thr-180 and Tyr-182 upon stimulation with RANKL in mouse bone marrow macrophages and pre osteoclasts. |
| CHUK | Protein | ncbigene:1147 | CHUK undergoes induced phosphorylation upon stimulation with RANKL in human umbilical vein endothelial cells (HUVECs) and RAW264.7 cells |
| MTOR | Protein | ncbigene:2475 | MTOR undergoes autophosphorylation upon stimulation with RANKL in osteoclasts and RAW264.7 murine macrophage cells. |
| SRC | Protein | ncbigene:6714 | SRC undergoes induced phosphorylation at tyrosine residue upon stimulation with RANKL in murine bone marrow derived macrophages, pre-osteoclasts and in RAW264.7 cells. |
| TNFSF11 | Protein | ncbigene:8600 | RANKL self associates to form a homotrimer. |
| TNFRSF11A | Protein | ncbigene:8792 | TNFRSF11A self assembled through its cytoplasmic domain at position 534-539 and formed a trimer in HEK293 cells transfected with TANFRF11A |
| TAB2 | Protein | ncbigene:23118 | |
| TAB1 | Protein | ncbigene:10454 | |
| RELA | Protein | ncbigene:5970 | |
| MAPK14 | Protein | ncbigene:1432 | |
| CBL | Protein | ncbigene:867 | RANKL stimulation induced the phosphorylation of CBL at Tyr-731 in mouse macrophage-like cell line, RAW264.7 |
| CDC42 | Protein | ncbigene:998 | |
| SQSTM1 | Protein | ncbigene:8878 | TRAF6 undergoes polyubiquitination upon stimulation with RANKL which is further enhanced by IFN-gamma in mouse macrophage cell line RAW 264.7 and in mouse bone marrow derived macrophages. |
| SPI1 | Protein | ncbigene:6688 | |
| MAP3K14 | Protein | ncbigene:9060 | RANKL stimulation leads to the activation of alternative NF-kappa B pathway which induced the formation of p52/RELB heterodimers through the activation of MAP3K14 (NIK). NIK deficient mice impairs RANKL mediated osteoclastogenesis. |
| NFKB2 | Protein | ncbigene:4791 | RANKL stimulation induces NFKB2 phosphorylation at Ser-866, Ser-870 in RAW264.7 cells. |
| NFKB2 | Protein | ncbigene:4791 |