Osteoblast differentiation and related diseases (WP4787)
Homo sapiens

{kind=link}
{kind=link}
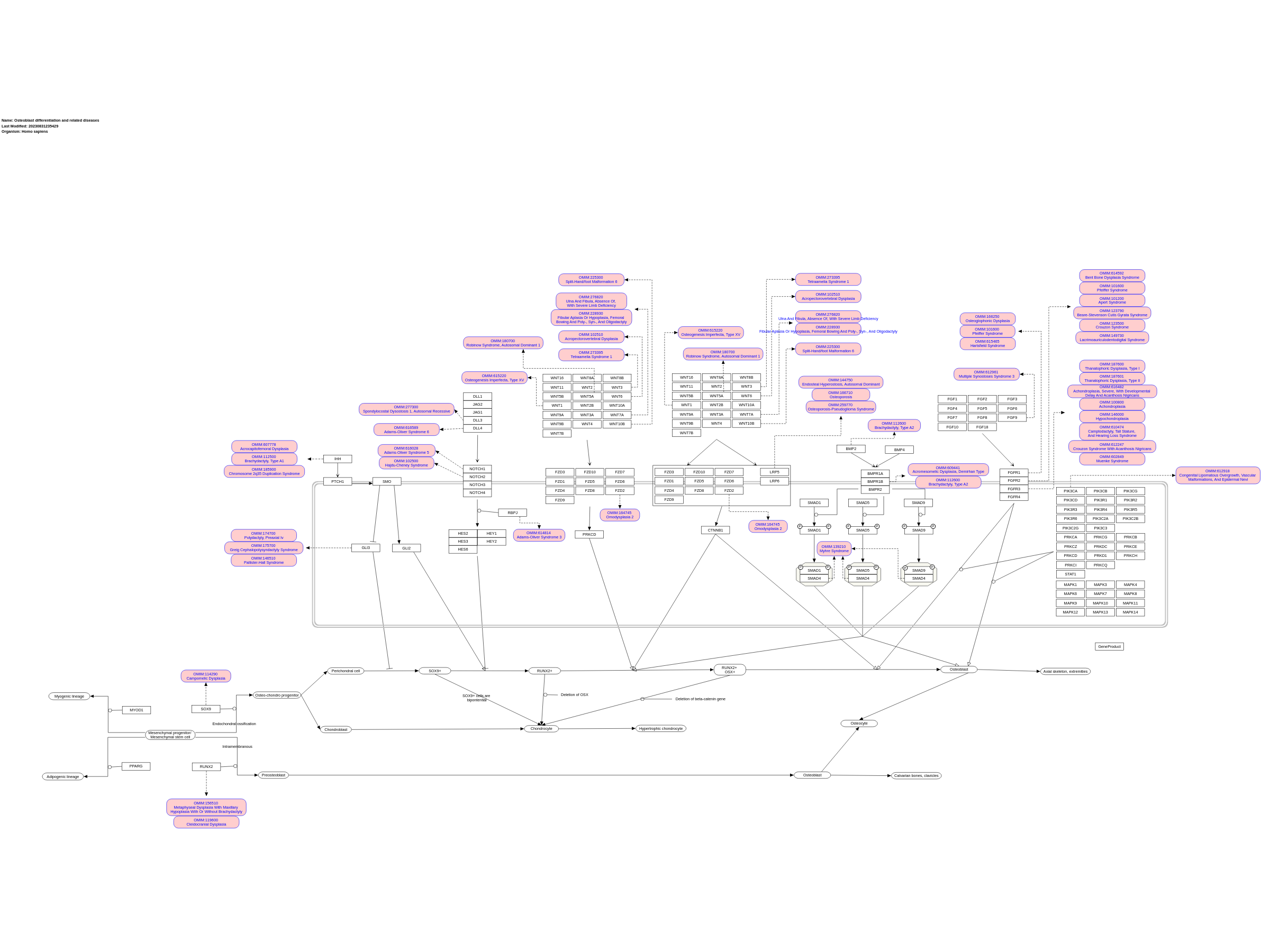
Taken from: Building stronger bones: molecular regulation of the osteoblast lineage by Fanxin Long. Osteoblast differentiation at a glance, Arkady Rutkovskiy, Kåre-Olav Stensløkken, Ingvar Jarle Vaage. Development of the endochondral skeleton by Fanxin Long, David Ornitz. Mesenchymal stem cells can give rise to 4 lineages by expressing corresponding transcriptional regulators: PPARg for adipogenic, MyoD for myogenic, Runx2 for osteoblastic, and Sox9 for chondrocytic lineages. In intramembranous ossification (osteogenesis in the scull and clavicles), preosteoblasts stem directly from mesenchymal stem cells, while in endochondral (osteogenesis of the axial skeleton and the limbs) a common osteo-chondro progenitor gives rise to both cell types. Extracellular signals regulating osteoblast differentiation: Model is based on studies of the mouse limb skeleton. Osteoblasts differentiate from mesenchymal progenitors (MP) through distinct developmental stages marked by expression of key transcription factors including SOX9, RUNX2, and OSX. Mature osteoblasts (OB) can further differentiate to osteocytes (OCY) or bone lining cells (not depicted) or undergo apoptosis (not depicted). Indian hedgehog (IHH) signaling is required for osteoblast differentiation during endochondral bone development. IHH binding to the receptor Patched homolog 1 (PTCH1) activates signaling through Smoothened (SMO), thereby inhibiting the generation of the proteolytically cleaved GLI3 repressor (GLI3R) and promoting the generation of the full-length GLI2 activator (GLI2A). Whereas derepression of GLI3R is sufficient to generate RUNX2+ cells, both derepression of GLI3R and activation of GLI2A are necessary for progression to the RUNX2+OSX+ stage. NOTCH signaling inhibits osteoblast differentiation. Following binding to their ligands, Jagged (JAG) or Delta-like (DLL), Notch receptors are proteolytically cleaved by the g-secretase complex, leading to release of the Notch intracellular domain (NICD) from the plasma membrane. NICD interacts with RBPJk and together they activate downstream target genes, including HES (Hairy and Enhancer of Split) and HEY (HES-related with YRPW motif) family transcription factors, ultimately leading to inhibition of osteoblast differentiation, seemingly at a stage before OSX activation. WNT signaling promotes osteoblast differentiation. During b-catenin-dependent WNT signaling, b-catenin is stabilized following binding of WNT to its receptors Frizzled (FZD) and lipoprotein receptor-related protein 5 (LRP5) or LRP6, leading to the transcription of b-catenin target genes and ultimately stimulating progression from the RUNX2+ stage to the RUNX2+OSX+ stage, and from RUNX2+OSX+ cells to mature osteoblasts. WNTcan also signal independently of LRP5/6 and b-catenin through protein kinase Cd (PKCd), promoting progression to the RUNX2+OSX+ stage through an unknown mechanism. Bone morphogenetic protein (BMP) signaling stimulates osteoblast differentiation and function. Binding of BMP2 or BMP4 to their receptors results in phosphorylation of SMAD1, SMAD5, or SMAD8. These can then form a complex with their partner, SMAD4, and enter the nucleus to regulate gene expression, ultimately promoting the transition to RUNX2+OSX+ cells and enhancing the function of mature osteoblasts; however, a direct role for SMAD signaling in osteoblast differentiation remains to be shown. Fibroblast growth factor (FGF) signaling has diverse roles in osteoblast lineage cells. FGFs function by binding to cell surface Tyr kinase FGF receptors (FGFR1–FGFR4 in humans and mice), leading to the activation of multiple signaling modules. FGF signaling regulates preosteoblast proliferation and osteoblast differentiation, as well as the function of mature osteoblasts. However, the precise stages at which FGFs regulate proliferation and differentiation, and the intracellular signaling cascades responsible for each function, remain to be elucidated. BMPR, BMP receptor; MAPK, mitogen-activated protein kinase; PI3K, phosphoinositide 3-kinase; STAT1, signal transducer and activator of transcription 1. Linked with a dotted arrow to the GeneProduct nodes are diseases caused by mutation in the respective gene. Protein phosphorylation sites were added based on information from PhosphoSitePlus (R), www.phosphosite.org.
For a description of pathway objects, see the WikiPathways Legend.
Authors
Ritchie Lee , Kristina Hanspers , and Eric WeitzActivity

Discuss this pathway
Check for ongoing discussions or start your own.
Cited In
Are you planning to include this pathway in your next publication? See How to Cite and add a link here to your paper once it's online.
Organisms
Homo sapiensCommunities
Skeletal DysplasiaAnnotations
Disease Ontology
Robinow syndrome Adams-Oliver syndrome brachydactyly type A1 osteogenesis imperfecta syndactyly type 1 campomelic dysplasia autosomal dominant Robinow syndrome 1 brachydactyly split hand-foot malformation 6 polydactyly cleidocranial dysplasia acrocapitofemoral dysplasia Pallister-Hall syndrome bone diseaseCell Type Ontology
osteoblast osteocyte chondrocytePathway Ontology
bone disease pathwayReferences
- Building strong bones: molecular regulation of the osteoblast lineage. Long F. Nat Rev Mol Cell Biol. 2011 Dec 22;13(1):27–38. PubMed Europe PMC Scholia
- Development of the endochondral skeleton. Long F, Ornitz DM. Cold Spring Harb Perspect Biol. 2013 Jan 1;5(1):a008334. PubMed Europe PMC Scholia
- PhosphoSitePlus, 2014: mutations, PTMs and recalibrations. Hornbeck PV, Zhang B, Murray B, Kornhauser JM, Latham V, Skrzypek E. Nucleic Acids Res. 2015 Jan;43(Database issue):D512-20. PubMed Europe PMC Scholia
- Osteoblast Differentiation at a Glance. Rutkovskiy A, Stensløkken KO, Vaage IJ. Med Sci Monit Basic Res. 2016 Sep 26;22:95–106. PubMed Europe PMC Scholia