NF-kB signaling and ARTD family members (WP5527)
Homo sapiens

{kind=link}
{kind=link}
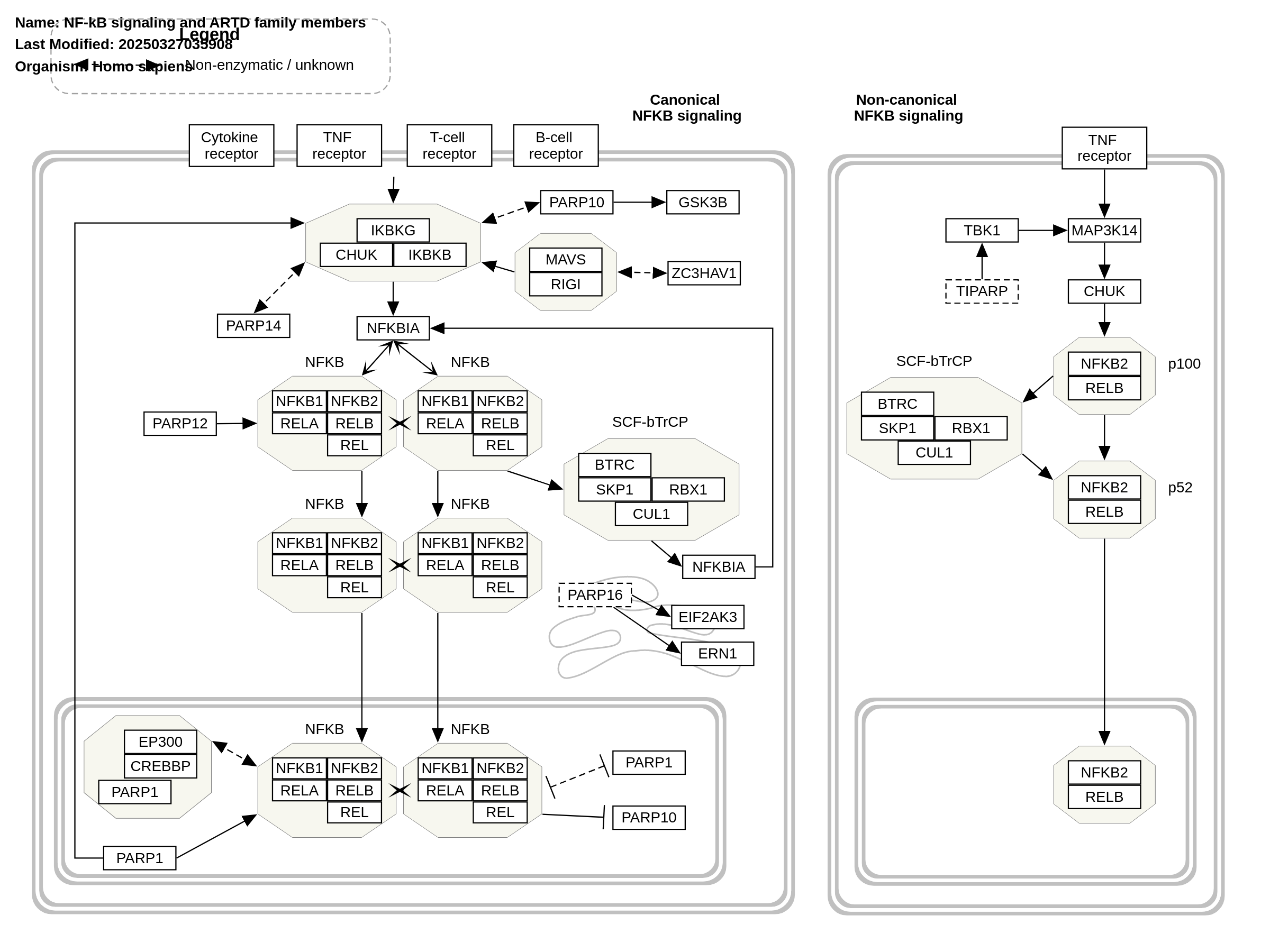
NF-κB binds to and represses the PARP10 promoter, resulting in transcriptional suppression in hepatocellular carcinoma. In prostate carcinoma resistant to gemcitabine, constitutive activation of NF-κB signaling is crucial, and elevated levels of PARP14 (ARTD8) are associated with poor patient outcomes. PARP14 plays a key role in cell survival and is essential for sustaining NF-κB signaling. Mechanistically, the reduction in NF-κB signaling is attributed to decreased phosphorylation of IκBα. The canonical NF-κB pathway is activated in response to viral infections. Mechanistically, the binding of 5′-triphosphate-modified RNA to RIG-I complexes enhances its interaction with MAVS, activating the IKK complex, which leads to IκBα degradation and the release of NF-κB dimers. Interestingly, the catalytically inactive RNA-binding protein PARP13 (ARTD13) restricts the replication of oncogenic viruses by stabilizing the activated RIG-I complex’s binding to MAVS, thus enhancing NF-κB signaling. This suggests that PARP13 may play a protective role against malignant transformation and cancer progression. PARP12 localizes to p62/SQSTM1 foci, and its activity is essential for initiating the NF-κB signaling cascade. ER-stress-induced inflammation and activation of the unfolded protein response (UPR) are transmitted through ER-associated stress sensors. These sensors significantly contribute to tumor progression and metastasis in an NF-κB-dependent manner. Mechanistically, activated IRE-1α recruits TRAF2, which activates the canonical pathway via IKK, leading to the release of NF-κB dimers. Simultaneously, PERK triggers an eIF2α-dependent reduction in translation, lowering overall IκB levels and shifting the equilibrium from complexed, inactive NF-κB to a free, transiently active form. During ER stress, PARP16, an ER-anchored protein, modifies itself and two stress sensors, IRE-1α and PERK. Activation of PARP16 promotes IRE-1α and PERK signaling, even in the absence of ER stress, and is crucial for their activity during the UPR. Both PARP1 (ARTD1) and its enzymatic activity are linked to increased NF-κB signaling. In TNFα-stimulated NIH3T3 fibroblasts lacking PARP1, NF-κB target gene expression is suppressed, despite NF-κB being localized in the nucleus. Moreover, LPS treatment of wild-type or PARP1-deficient mice reinforces the involvement of PARP1 in NF-κB’s transcriptional activity. Mechanistically, LPS-dependent NF-κB activation in primary murine fibroblasts is driven by the interaction of PARP1 with transcriptional coactivators CREB-binding protein (CBP) and p300. This complex formation leads to PARP1 acetylation, enabling the PARP1-CBP-p300 complex to interact with the p50 subunit of NF-κB, thus activating NF-κB signaling and initiating the transcription of proinflammatory cytokines, chemokines, transcription factors, and other inflammatory mediators. In non-canonical NF-κB signaling, NF-κB2 processing is regulated by NIK, which is downregulated by non-canonical IKK TBK1. In an NF-κB-independent context, TBK1 activates IRF3 in response to viral infections. Notably, TBK1 kinase activity and antiviral response efficacy are negatively regulated by PARP7 (ARTD14/TIPARP)-mediated ADP-ribosylation. In HeLa and U2OS cells, PARP10 inhibits IKK complex activation and NF-κB signaling. Mechanistically, PARP10 reduces K63-linked polyubiquitination of NEMO, which prevents IKK complex assembly and activation. Inspired by Figure 1 and associated description in Boehi et al. (2021).
For a description of pathway objects, see the WikiPathways Legend.
Authors
Eric WeitzActivity

Discuss this pathway
Check for ongoing discussions or start your own.
Cited In
Are you planning to include this pathway in your next publication? See How to Cite and add a link here to your paper once it's online.
Organisms
Homo sapiensCommunities
Annotations
Disease Ontology
hepatocellular carcinoma prostate carcinoma viral infectious diseasePathway Ontology
nuclear factor kappa B signaling pathway| Label | Type | Compact URI | Comment |
|---|---|---|---|
| IKBKG | GeneProduct | ensembl:ENSG00000269335 | 'NEMO' in source |
| CHUK | GeneProduct | ensembl:ENSG00000213341 | 'IKK1' in source |
| IKBKB | GeneProduct | ensembl:ENSG00000104365 | 'IKK2' in source |
| NFKBIA | GeneProduct | ensembl:ENSG00000100906 | 'IKB' in source |
| EIF2AK3 | GeneProduct | ensembl:ENSG00000172071 | 'PERK' in source |
| ERN1 | GeneProduct | ensembl:ENSG00000178607 | 'IRE-1a' in source |
| EP300 | GeneProduct | ensembl:ENSG00000100393 | 'p300' in source |
| CREBBP | GeneProduct | ensembl:ENSG00000005339 | 'CBP' in source |
| PARP10 | GeneProduct | ensembl:ENSG00000178685 | |
| GSK3B | GeneProduct | ensembl:ENSG00000082701 | |
| MAVS | GeneProduct | ensembl:ENSG00000088888 | |
| RIGI | GeneProduct | ensembl:ENSG00000107201 | 'RIG-I' in source |
| ZC3HAV1 | GeneProduct | ensembl:ENSG00000105939 | 'PARP13' in source |
| PARP14 | GeneProduct | ensembl:ENSG00000173193 | |
| PARP12 | GeneProduct | ensembl:ENSG00000059378 | |
| PARP1 | GeneProduct | ensembl:ENSG00000143799 | |
| TIPARP | GeneProduct | ensembl:ENSG00000163659 | 'PARP7' in source |
| TBK1 | GeneProduct | ensembl:ENSG00000183735 | |
| MAP3K14 | GeneProduct | ensembl:ENSG00000006062 | 'NIK' in source |
| NFKB2 | GeneProduct | ensembl:ENSG00000077150 | 'P100' in source |
| RELB | GeneProduct | ensembl:ENSG00000104856 | |
| NFKB2 | GeneProduct | ensembl:ENSG00000077150 | 'P52' in source |
| RBX1 | GeneProduct | ensembl:ENSG00000100387 | |
| SKP1 | GeneProduct | ensembl:ENSG00000113558 | |
| CUL1 | GeneProduct | ensembl:ENSG00000055130 | |
| BTRC | GeneProduct | ensembl:ENSG00000166167 | |
| NFKB1 | GeneProduct | ensembl:ENSG00000114515 | |
| RELA | GeneProduct | ensembl:ENSG00000184697 | |
| NFKB2 | GeneProduct | ensembl:ENSG00000114526 | |
| RELB | GeneProduct | ensembl:ENSG00000103171 | |
| REL | GeneProduct | ensembl:ENSG00000105825 | |
| PARP16 | GeneProduct | ensembl:ENSG00000138617 |