Sphingolipid metabolism in senescence (WP5121)
Homo sapiens

{kind=link}
{kind=link}
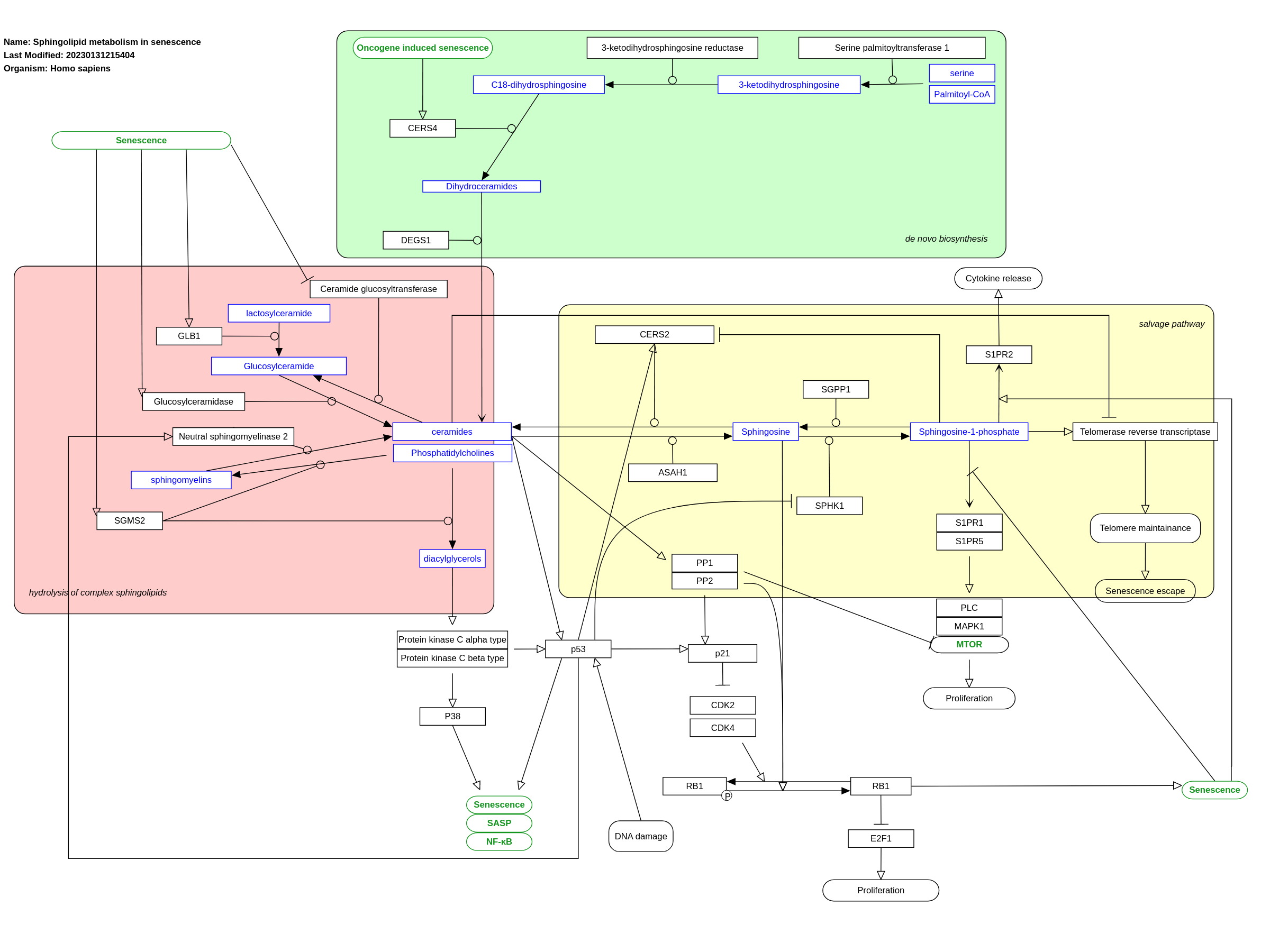
Sphingolipids, which include sphingomyelins, sphingosines and ceramides are bioactive bioactive molecules present in all eukaryotic cells, are important in regulating various aspects of cell biology, such as cell cycle, proliferation, and even senescence (Trayssac et al., 2018). Evidence shows that ceramide metabolism is deregulated in cellular senescence and can even induce it. (Venable et al., 2006). Ceramides are thought to induce senescence in a p53 dependent and independent manner by hypophosphorylated retinoblastoma protein (Lee et al., 2000; Dix 2018; Jeffries & Krupenko, 2018). Ceramides function both down- and upstream of p53 (Jeffries et al., 2018). p53 mediates the interference of the phosphorylation of retinoblastoma-like protein RBL1 and RBL through the cyclin-dependent kinase inhibitor p21 (Jeffries et & Krupenko, 2018). Additionally, p53 has been demonstrated to directly activate ceramide synthase. (Jeffries & Krupenko, 2018). Consequently, this implies a feedback mechanism between ceramides and the tumor suppressor. Ceramide also interacts directly with human telomerase reverse transcriptase (hTERT) by inhibiting it, therefore causing telomere instability (Hannun & Obeid, 2002; Deevska et al., 2021). Ceramides activate protein phosphatase 1 and 2A (PP1 and PP2A), which increases the levels of p21 (Trayssac et al., 2018). This then inhibits the cyclin dependent kinase 2 (CDK2) and CDK4. As a consequence, the retinoblastoma protein is hypophosphorylated and induces senescence (Lee et al., 2000; Dix 2018). The hypophosphorylated retinoblastoma protein leads to the inhibition of E2F (Dix, 2018; Jeffries & Krupenko, 2018). This group of genes is normally responsible for cell proliferation and therefore their inhibition causes senescence (Dix 2018). In addition to that, PP1 and PP2A directly interfere with Rb by dephosphorylating it (Dix 2018). These two phosphatases also inhibit the mTOR pathway that is associated with cell proliferation (Millner & Atilla-Gokcumen, 2020). Sphingosine-1-phosphate (S1P) is thought to induce cell proliferation and migration by binding to S1PR1 and S1PR5. This is downregulated in senescence (Trayssac et al., 2018). In cellular senescence, S1P has been shown to be depleted as a result of a downregulation of sphingosine kinase 1 (SPHK1), induced by p53. The downregulation of SK1 is thought to be due to its degradation induced by p53 (Kim et al., 2019). Moreover, S1P has hTERT as a direct target and promotes its stability (Magali et al., 2021). Furthermore, there is an increase in S1P binding to the S1P receptor 2 (S1PR2), which has been associated with the release of pro-inflammatory cytokines and therefore the SASP. S1P is also thought to inhibit ceramide synthase 2 (CERS2), which catalyzes the conversion of sphingosine into ceramide (Magali et al., 2021). Sphingosine is increased in senescence cells by the increase of S1P-phopshatase (SGPP1) and the increase in acid ceramidase ASAH-1 SGPP1 catalyzes the conversion of S1P to sphingosine, while ASAH-1 catalyzes the conversion of ceramide to sphingosine (Munk et al., 2021; Kim et al., 2019). This causes the dephosphorylation of retinoblastoma protein, which further causes senescence (Trayssac et al., 2018). Ceramide can be synthesized de novo starting with the conversion of palmitoyl-CoA and serine to 3-ketodihydrosphingosine. Upregulation of ceramide synthase 4 (CERS4), which converts dihydro-sphingosine into dihydro-ceramide. CERS4 has been thought to be a key enzyme in two types of senescence: OIS and replicative senescence, by mediating its effects through the PP1-Rb-E2F axis (Dix, 2018) Galactosidase β1 (GLB1) and in glucosylceramidase (GBA), which catalyze the conversion of lactosylceramide to glucosylceramide and the conversion of glucosylceramide to ceramide, respectively, are upregulated in cellular senescence. Additionally, there is a decrease in glucosylceramidase synthase, which catalyzes the formation of glucosylceramide from ceramide (Flor et al., 2017). Activation of p53, due to DNA damage, leads to an increase of neutral sphingomyelinase 2 (nSMase 2) (Deevska et al., 2021; Jeffries & Krupenko, 2018). Moreover, it has been noted that in senescent cells, there are elevated levels of neutral sphingomyelinases ( Millner & Atilla-Gokcumen, 2020). These enzymes catalyze the conversion of sphingomyelin to ceramide (Gey & Seeger, 2013). Elevated ceramide levels result in elevated diacylglycerol (DAG) levels (Deevska et al., 2021). These two lipids are linked by sphingomyelin synthase 2 (SGMS2). SGMS2 catalyzes the transfer of a phosphorycholine group from phosphatidylcholine to ceramide, yielding sphingomyelin and DAG. In cellular senescence, SMS was found to be overexpressed. This caused an increase in DAG which caused the chronic activation of its downstream effectors. This causes the downregulation of protein kinase C α and β (PKCα and β) via the prolonged activation of phospholipase C. The consequences of this is the activation of p53 and p38, leading to SASP and senescence (Deevska et al., 2021).
For a description of pathway objects, see the WikiPathways Legend.
Authors
Tade Idowu , Denise Kalbe , Mario Zubak , Doreen Jost , Eric Weitz , and Lars WillighagenActivity

Discuss this pathway
Check for ongoing discussions or start your own.
Cited In
Are you planning to include this pathway in your next publication? See How to Cite and add a link here to your paper once it's online.
Organisms
Homo sapiensCommunities
Annotations
Pathway Ontology
aging pathway sphingolipid metabolic pathway| Label | Type | Compact URI | Comment |
|---|---|---|---|
| 3-ketodihydrosphingosine | Metabolite | chebi:17862 | |
| lactosylceramide | Metabolite | chebi:134507 | |
| Sphingosine-1-phosphate | Metabolite | chebi:37550 | |
| C18-dihydrosphingosine | Metabolite | chebi:16566 | |
| Glucosylceramide | Metabolite | chebi:18368 | |
| ceramides | Metabolite | chebi:17761 | |
| Phosphatidylcholines | Metabolite | chebi:49183 | |
| Dihydroceramides | Metabolite | chebi:139048 | |
| p53 | Metabolite | chebi:77731 | |
| diacylglycerols | Metabolite | chebi:18035 | |
| sphingomyelins | Metabolite | chebi:62490 | |
| serine | Metabolite | chebi:17822 | |
| Sphingosine | Metabolite | chebi:45719 | |
| Palmitoyl-CoA | Metabolite | chebi:15525 | |
| CDK4 | GeneProduct | ensembl:ENSG00000135446 | |
| P38 | GeneProduct | ensembl:ENSG00000152464 | |
| SPHK1 | GeneProduct | ensembl:ENSG00000176170 | |
| RB1 | GeneProduct | ensembl:ENSG00000139687 | |
| S1PR5 | GeneProduct | ensembl:ENSG00000180739 | |
| S1PR2 | GeneProduct | ensembl:ENSG00000267534 | |
| S1PR1 | GeneProduct | ensembl:ENSG00000170989 | |
| CERS4 | GeneProduct | ensembl:ENSG00000090661 | |
| SGMS2 | GeneProduct | ensembl:ENSG00000164023 | |
| Neutral sphingomyelinase 2 | GeneProduct | ensembl:ENSG00000103056 | |
| SGPP1 | GeneProduct | ensembl:ENSG00000126821 | |
| MTOR | GeneProduct | ensembl:ENSG00000198793 | |
| CERS2 | GeneProduct | ensembl:ENSG00000143418 | |
| DEGS1 | GeneProduct | ensembl:ENSG00000143753 | |
| MAPK1 | GeneProduct | ncbigene:5594 | |
| CDK2 | GeneProduct | ensembl:ENSG00000123374 | |
| E2F1 | GeneProduct | ensembl:ENSG00000101412 | |
| ASAH1 | GeneProduct | ensembl:ENSG00000104763 | |
| GLB1 | GeneProduct | ensembl:ENSG00000170266 | |
| Telomerase reverse transcriptase | Protein | uniprot:O14746 | |
| Protein kinase C alpha type | Protein | uniprot:P17252 | |
| Protein kinase C beta type | Protein | uniprot:P05771 | |
| 3-ketodihydrosphingosine reductase | Protein | uniprot:Q06136 | |
| PP1 | Protein | wikidata:Q27088427 | |
| PP2 | Protein | wikidata:Q7120082 | |
| Glucosylceramidase | Protein | uniprot:A0A068F658 | |
| p21 | Protein | uniprot:A0A024RCX5 | |
| Ceramide glucosyltransferase | Protein | uniprot:Q16739 | |
| Serine palmitoyltransferase 1 | Protein | uniprot:O15269 | |
| PLC | Protein | uniprot:A0A087WT80 |
References
- The Ceramide-centric universe of lipid-mediated cell regulation: stress encounters of the lipid kind. Hannun YA, Obeid LM. J Biol Chem. 2002 Jul 19;277(29):25847–50. PubMed Europe PMC Scholia
- An overview of sphingolipid metabolism: from synthesis to breakdown. Gault CR, Obeid LM, Hannun YA. Adv Exp Med Biol. 2010;688:1–23. PubMed Europe PMC Scholia
- Metabolic Changes Investigated by Proton NMR Spectroscopy in Cells Undergoing Oncogene-Induced Senescence. Gey C, Seeger K. Methods Mol Biol. 2017;1534:155–63. PubMed Europe PMC Scholia
- A signature of enhanced lipid metabolism, lipid peroxidation and aldehyde stress in therapy-induced senescence. Flor AC, Wolfgeher D, Wu D, Kron SJ. Cell Death Discov. 2017 Oct 30;3:17075. PubMed Europe PMC Scholia
- Ceramide Signaling and p53 Pathways. Jeffries KA, Krupenko NI. Adv Cancer Res. 2018;140:191–215. PubMed Europe PMC Scholia
- Role of sphingolipids in senescence: implication in aging and age-related diseases. Trayssac M, Hannun YA, Obeid LM. J Clin Invest. 2018 Jul 2;128(7):2702–12. PubMed Europe PMC Scholia
- Links between accelerated replicative cellular senescence and down-regulation of SPHK1 transcription. Kim MK, Lee W, Yoon GH, Chang EJ, Choi SC, Kim SW. BMB Rep. 2019 Mar;52(3):220–5. PubMed Europe PMC Scholia
- Lipid Players of Cellular Senescence. Millner A, Atilla-Gokcumen GE. Metabolites. 2020 Aug 21;10(9):339. PubMed Europe PMC Scholia
- Targeting sphingosine kinase 1 (SK1) enhances oncogene-induced senescence through ceramide synthase 2 (CerS2)-mediated generation of very-long-chain ceramides. Trayssac M, Clarke CJ, Stith JL, Snider JM, Newen N, Gault CR, et al. Cell Death Dis. 2021 Jan 4;12(1):27. PubMed Europe PMC Scholia
- Onset of Senescence and Steatosis in Hepatocytes as a Consequence of a Shift in the Diacylglycerol/Ceramide Balance at the Plasma Membrane. Deevska G, Dotson PP 2nd, Mitov M, Butterfield DA, Nikolova-Karakashian M. Cells. 2021 May 21;10(6):1278. PubMed Europe PMC Scholia
- Acid ceramidase promotes senescent cell survival. Munk R, Anerillas C, Rossi M, Tsitsipatis D, Martindale JL, Herman AB, et al. Aging (Albany NY). 2021 Jun 8;13(12):15750–69. PubMed Europe PMC Scholia